Imperial celebrates Kuwait links with first alumni event
Superfast physics and a trio of Fellows: News from Imperial
'Living paint’ startup wins Imperial’s top entrepreneurship prize
Our organisation comprises eight world-class departments spread over seven West London campuses
The mission of the Department of Brain Sciences is to develop and sustain a unique portfolio of translational research in neuroscience and mental health that is recognised internationally for scientific excellence.
Find out more
The Department of Immunology and Inflammation’s mission is to integrate discovery science with clinical medicine to prevent or treat inflammatory and haematological conditions. We focus on discovering fundamental mechanisms linking the immune system to disease and translating these to advance molecular medicine in the clinic.
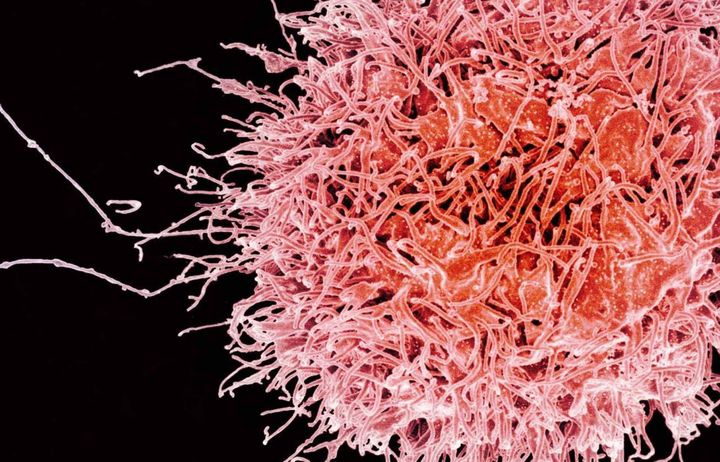
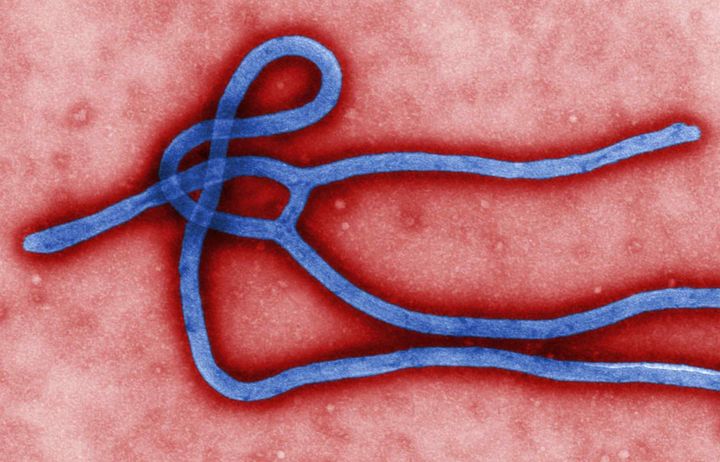
The new Department of Infectious Disease aims to understand the role of microbes in health and disease and to describe host responses to pathogens, both natural and those induced by vaccines, at molecular, structural and systems levels.
The Institute of Clinical Sciences' focus is on fundamental research to explore the interface between genomics, physiology and epigenetics to gain a better understanding of metabolic diseases and how best to treat them.
The Department of Metabolism, Digestion and Reproduction encompasses a broad spectrum of clinical specialties and academic disciplines across Reproductive & Developmental Biology; Digestive Diseases; Diabetes, Endocrinology & Metabolism; and Systems Medicine.
The NHLI is one of the largest heart and lung departments in the world and retains its position as a highly respected research centre carrying out research, development and education in cardiovascular and respiratory science.
The School of Public Health aims to achieve better health in the population through strengthening the public health science base, training the next generation of public health leaders and influencing health policies around the world.
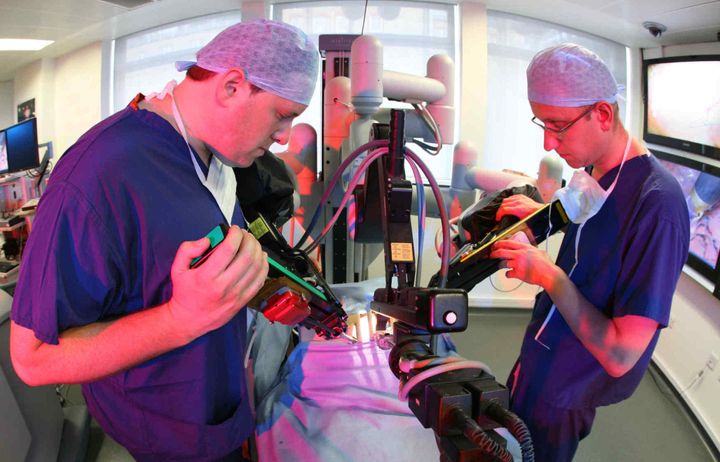
The Department of Surgery and Cancer aims to innovate, develop and apply new technologies, techniques and computationally intense systems medicine approaches to enhance medical and surgical diagnostics and prognostics.