





Key areas


Spotlight on postgraduate programmes
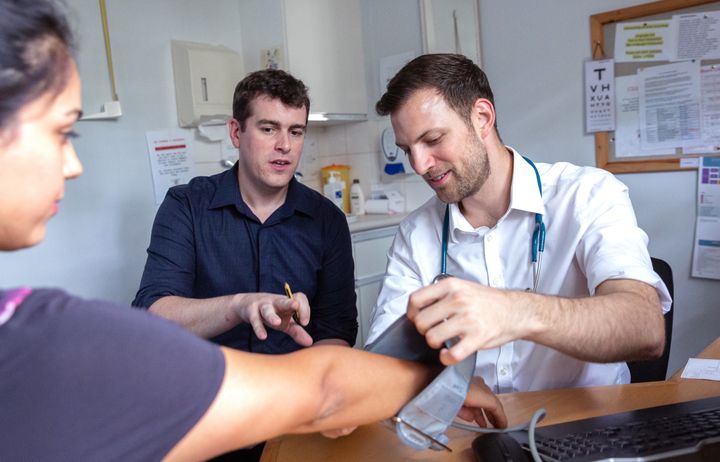
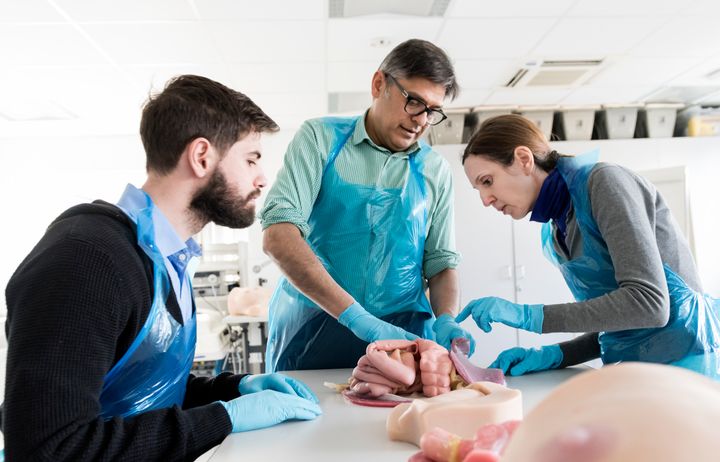
Our Master’s programmes draw on recognised excellence in research to offer a wide range of programmes for careers in lab-based science, public health and clinical healthcare.
Our Master’s programmes draw on recognised excellence in research to offer a wide range of programmes for careers in lab-based science, public health and clinical healthcare.



